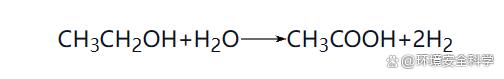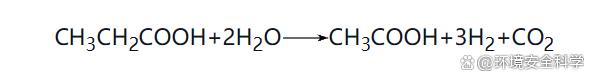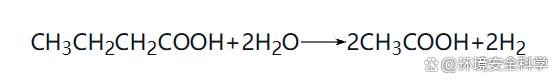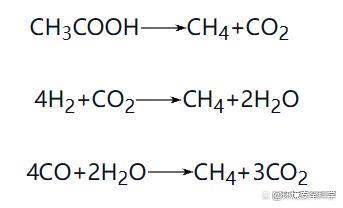當(dāng)前位置:首頁>生活> 污泥的厭氧消化微生物學(xué)原理
發(fā)布時(shí)間:2025-10-28閱讀( 20)
|
在厭氧消化過程中,參與反應(yīng)的細(xì)菌分為兩大類:一類是產(chǎn)氫產(chǎn)乙酸菌、水解酸化菌和同型產(chǎn)乙酸菌等非產(chǎn)甲烷菌;另一類是產(chǎn)甲烷菌。由于微生物種群之間的生態(tài)平衡關(guān)系是支配厭氧消化過程的本質(zhì)規(guī)律,所以在本節(jié)中著重介紹厭氧消化過程微生物的種類、特性及其相互代謝關(guān)系。 (1)厭氧消化過程微生物種類 1)非產(chǎn)甲烷菌 ① 水解發(fā)酵菌(產(chǎn)酸細(xì)菌)。主要的發(fā)酵產(chǎn)酸細(xì)菌包括梭菌屬、擬桿菌屬、丁酸弧菌屬、雙歧桿菌屬等。其主要功能是水解和酸化作用。水解——在胞外酶的作用下,將不溶性大分子有機(jī)物水解成可溶性有機(jī)物;酸化——將可溶性大分子有機(jī)物轉(zhuǎn)化為乙酸、丙酸、丁酸、戊酸等有機(jī)酸及醇類等。水解發(fā)酵細(xì)菌的水解過程較緩慢,并受SRT、pH值、有機(jī)物種類等多種因素的影響,但產(chǎn)酸反應(yīng)的速率較快。發(fā)酵產(chǎn)酸細(xì)菌可以按功能分為蛋白質(zhì)分解菌、半纖維素分解菌、淀粉分解菌、纖維素分解菌、脂肪分解菌等。 ② 產(chǎn)氫產(chǎn)乙酸菌。該類細(xì)菌的主要功能是將各種高級(jí)脂肪酸和醇類氧化分解為乙酸、CO2和H2,為產(chǎn)甲烷階段提供合適的基質(zhì),在厭氧系統(tǒng)中常常與產(chǎn)甲烷細(xì)菌處于共生互營關(guān)系。主要的產(chǎn)氫產(chǎn)乙酸反應(yīng)有:
只有在乙酸濃度和系統(tǒng)中氫分壓很低時(shí)上述反應(yīng)才能順利進(jìn)行,因此,產(chǎn)氫產(chǎn)乙酸反應(yīng)的順利進(jìn)行,常常需要后續(xù)產(chǎn)甲烷反應(yīng)能及時(shí)將其主要的兩種產(chǎn)物乙酸和H2消耗掉。主要的產(chǎn)氫產(chǎn)乙酸細(xì)菌多數(shù)是專性厭氧菌和兼性厭氧菌,包括互營桿菌屬、互營單胞菌屬、梭菌屬、暗桿菌屬等。 ③ 同型產(chǎn)乙酸菌。在“三階段理論”的影響下,人們將注意力集中在產(chǎn)氫產(chǎn)乙酸菌與產(chǎn)甲烷菌等的互營作用上,并將其歸入產(chǎn)甲烷相進(jìn)行研究,卻忽略了產(chǎn)酸菌、產(chǎn)氫產(chǎn)乙酸菌與同型乙酸菌之間的相互作用。值得一提的是,同型產(chǎn)乙酸菌在厭氧產(chǎn)酸過程中,具有特殊作用,它可以將二氧化碳和氫氣轉(zhuǎn)化為乙酸,在氫分壓高時(shí),它的作用增強(qiáng);在氫分壓低時(shí),它的作用減弱,起到了一個(gè)平衡器的作用。但在一般厭氧消化器中,同型產(chǎn)乙酸菌所產(chǎn)的乙酸不到4%。盡管人們對(duì)自然生態(tài)系統(tǒng)中同型產(chǎn)乙酸菌對(duì)碳循環(huán)的重要貢獻(xiàn)有較深入的認(rèn)識(shí),但卻沒能在人工厭氧產(chǎn)酸系統(tǒng)中充分發(fā)揮同型產(chǎn)乙酸菌的作用。 2)產(chǎn)甲烷菌(Methanogens) 產(chǎn)甲烷菌是產(chǎn)甲烷階段的主要細(xì)菌,屬于嚴(yán)格的厭氧菌,主要代謝產(chǎn)物是甲烷。主要可分為兩大類:乙酸營養(yǎng)型產(chǎn)甲烷菌和H2營養(yǎng)型產(chǎn)甲烷菌。 產(chǎn)甲烷細(xì)菌的主要功能是將產(chǎn)氫產(chǎn)乙酸菌的產(chǎn)物——乙酸和H2/CO2轉(zhuǎn)化為CH4和CO2,使厭氧消化過程得以順利進(jìn)行。一般來說,在自然界中,乙酸營養(yǎng)型產(chǎn)甲烷菌的種類較少,只有產(chǎn)甲烷八疊球菌(Methanosarcina)和產(chǎn)甲烷絲狀菌(Methanothrix),但這兩種產(chǎn)甲烷細(xì)菌在厭氧反應(yīng)器中居多,特別是后者,因?yàn)樵趨捬醴磻?yīng)器中,乙酸是主要的產(chǎn)甲烷基質(zhì),一般來說,有70%左右的甲烷是來自乙酸的氧化分解。 典型的產(chǎn)甲烷反應(yīng)有:
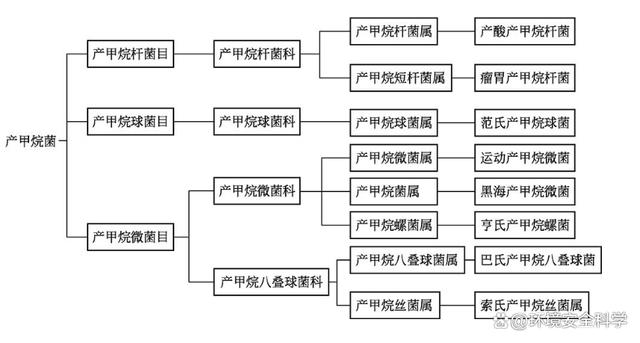
根據(jù)產(chǎn)甲烷菌的形態(tài)和生理生態(tài)特征,可將其進(jìn)行分類,如圖所示。
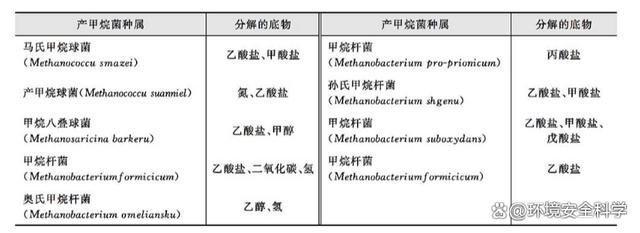
迄今為止,可分離鑒定的產(chǎn)甲烷菌達(dá)到70余種,分屬于3個(gè)目,7個(gè)科,19個(gè)屬。產(chǎn)甲烷菌有各種不同的形態(tài),我們常見的產(chǎn)甲烷菌有產(chǎn)甲烷桿菌、產(chǎn)甲烷球菌、產(chǎn)甲烷螺旋菌、產(chǎn)甲烷八疊球菌四類。下表列示了幾種主要產(chǎn)甲烷菌種屬及其分解的底物。
厭氧消化過程中產(chǎn)甲烷菌的生長情況直接關(guān)系到甲烷的產(chǎn)量,其能分解的物質(zhì)均是由大分子有機(jī)物分解生成的小分子中間產(chǎn)物,本身并沒有直接分解大分子有機(jī)物的能力。 產(chǎn)甲烷菌是通過細(xì)胞分裂進(jìn)行繁殖。下面兩種物質(zhì)可以大大促進(jìn)這一細(xì)胞分裂作用,有助于產(chǎn)甲烷菌的繁殖,但是,這二者之間又存在著相互排斥的作用。 ① 濾泡激素(follicular hormone,follikehoron):公馬或妊娠的母馬尿中得到的激素。 ② 細(xì)菌酶(bacteriozym):從酶細(xì)胞中得到的植物性提取物。 無論哪一種產(chǎn)甲烷菌的生存時(shí)間都不少于幾個(gè)星期,因此,一個(gè)新建或者一度停用的消化池,會(huì)需要很長的啟動(dòng)時(shí)間。但是,通過良好的消化污泥進(jìn)行定期接種,也可在短時(shí)間內(nèi)最大限度地發(fā)揮細(xì)菌的作用。 產(chǎn)甲烷菌由于具有某種性質(zhì)的生物排他性,因此,在產(chǎn)甲烷菌大量繁殖的地方,其他微生物的種類和數(shù)量就受到抑制。 細(xì)菌的生長和繁殖,通常是依靠同化作用和異化作用這兩種不同的代謝活動(dòng)來維持的。與其他植物一樣,細(xì)菌對(duì)養(yǎng)分的吸收與代謝產(chǎn)物的排出,是通過細(xì)胞膜的滲透作用進(jìn)行的。一般來講,細(xì)菌體內(nèi)的代謝產(chǎn)物濃度高于周圍液體的濃度時(shí),由于滲透壓的作用,細(xì)菌將從周圍吸收水分。但是,細(xì)胞膜彈性產(chǎn)生的滲透壓與反滲透壓達(dá)到平衡時(shí),細(xì)菌便膨大而處于正常狀態(tài)。但是,隨著代謝產(chǎn)物的增加,細(xì)菌體內(nèi)的濃度低于周圍液體的濃度時(shí),滲透壓則起反作用,從細(xì)菌體內(nèi)奪走水分,導(dǎo)致原生質(zhì)(protoplasm)萎縮,即使周圍還有水,細(xì)菌的生命也將就此終止,為促進(jìn)消化過程而進(jìn)行的攪拌就是要避免產(chǎn)生這種情況。只要存在養(yǎng)分并具備必要的生活條件,細(xì)菌就能不斷地進(jìn)行吸收養(yǎng)分與代謝作用。 對(duì)于產(chǎn)甲烷菌來講,最重要的養(yǎng)料是碳和氮,但其他有機(jī)物和無機(jī)物對(duì)其生存也有影響。有機(jī)的有效物質(zhì)是酶和維生素。酶大致可以分為以下幾種。 ① 氧化還原酶類 氧化還原酶類大致可以分為脫氫酶和氧化酶兩種。脫氫酶可以活化底物上的氫,并使它轉(zhuǎn)移到另一種物質(zhì)上,使底物因脫氫而氧化,不同的底物將由不同的脫氫酶進(jìn)行脫氫作用。氧化酶能將分子氧(空氣中的氧)活化,從而作為氫的受體而形成水或催化底物脫氫,并氧化生成過氧化氫。 ② 轉(zhuǎn)移酶類 轉(zhuǎn)移酶類能催化一種化合物分子的基團(tuán)轉(zhuǎn)移到另一種化合物分子上。 ③ 水解酶類 水解酶類能催化大分子有機(jī)物的水解作用及其逆反應(yīng)。 ④ 裂解酶類 裂解酶類能催化有機(jī)物碳鏈的斷裂,產(chǎn)生碳鏈較短的產(chǎn)物。 ⑤ 異構(gòu)酶類 異構(gòu)酶類能催化同分異構(gòu)化合物之間的相互轉(zhuǎn)化,即分子內(nèi)部基團(tuán)的重新排列。 ⑥ 合成酶類 合成酶類能催化有三磷酸腺苷參加的合成反應(yīng),這類酶關(guān)系著很多重要生命物質(zhì)的合成。 在無機(jī)的有效物質(zhì)中,在數(shù)量上比較主要的是磷酸(P2O5)和鉀(灰分)(K2O)。此外,鐵、鈉、鎂、銅、硫、鉬、鈷、錫、鈀等痕量元素對(duì)產(chǎn)甲烷菌的生長也有影響。少量的硫化氫有助于細(xì)菌繁殖,但濃度一高就成為有害物質(zhì)。 另外,必須創(chuàng)造一個(gè)弱堿環(huán)境,最佳pH值應(yīng)保持在7.0~7.5,有機(jī)酸濃度應(yīng)在2000~3000mg/L以下。這一容許濃度將根據(jù)氨和其他陽離子存在的情況稍有變動(dòng)。有機(jī)酸濃度是通過迪克洛(Duclaux)法以乙酸的含量測定的,其值與pH值無關(guān)。有機(jī)酸濃度如果在2000~3000mg/L以上,產(chǎn)甲烷菌的活動(dòng)能力將逐漸減弱,消化速度也慢了。可通過停止供給生污泥、降低污泥含固率、投入消石灰等方法予以解決;但是,如果適應(yīng)產(chǎn)甲烷菌生活的條件不能恢復(fù),污泥就將開始酸性發(fā)酵。當(dāng)有機(jī)酸濃度在5000mg/L以上,這些處理措施幾乎無效。另一方面,最佳堿度的上限,換算成CaCO3為2000mg/L左右。 在生物分類學(xué)上,產(chǎn)甲烷菌(Methanogens)屬于古細(xì)菌(Archaebacteria),大小、外觀上與普通細(xì)菌(Eubacteria)相似,但實(shí)際上,其細(xì)胞成分特殊,特別是細(xì)胞壁的結(jié)構(gòu)較特殊。產(chǎn)甲烷菌在自然界的分布,一般可以認(rèn)為是棲息于一些極端環(huán)境(如地?zé)崛⑸詈;鹕娇凇⒊练e物等)中,但實(shí)際上其分布極為廣泛,如污泥、瘤胃、昆蟲腸道、濕樹木、厭氧反應(yīng)器等。產(chǎn)甲烷菌是嚴(yán)格的厭氧細(xì)菌,氧和氧化劑對(duì)其有很強(qiáng)的毒害作用,要求氧化還原電位在-400~-150mV。產(chǎn)甲烷菌的增殖速率很慢,繁殖世代時(shí)間長,可達(dá)4~6d。產(chǎn)甲烷菌的生存溫度為0~80℃,低溫、中溫、高溫細(xì)菌的最佳溫度分別為15℃、30℃、55℃左右。與兼性厭氧菌比較,產(chǎn)甲烷菌更為敏感。但是,如果給產(chǎn)甲烷菌創(chuàng)造了最佳生活條件,就會(huì)得到良好的消化污泥。
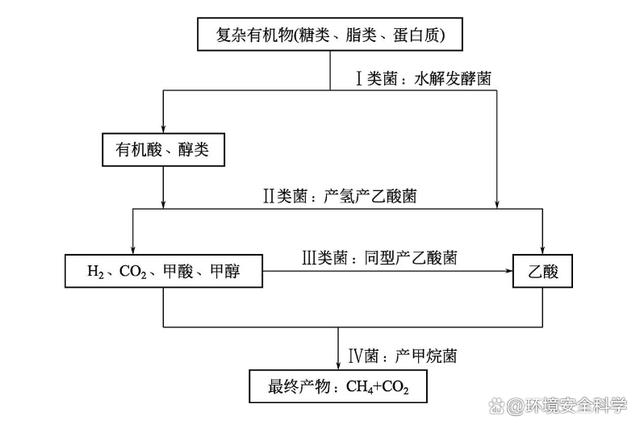
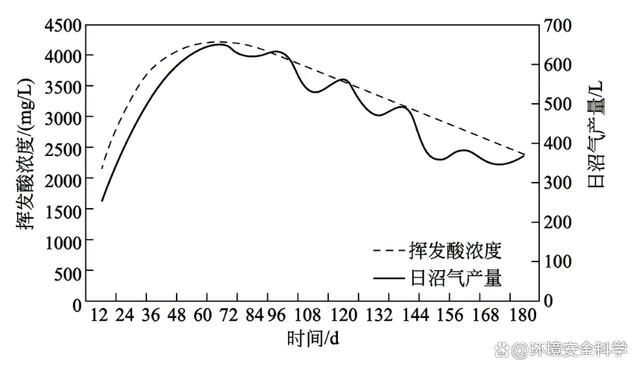
(2)厭氧消化過程中微生物的代謝關(guān)系 厭氧消化是一個(gè)多菌群共同代謝的過程,各類菌群之間的代謝作用相輔相成。在整個(gè)厭氧代謝過程中,產(chǎn)甲烷菌并不是孤立進(jìn)行的,它周圍眾多的菌群先于產(chǎn)甲烷菌代謝,以此提供產(chǎn)甲烷菌正常代謝的條件。從物質(zhì)代謝轉(zhuǎn)化的角度來看,首先,有機(jī)物的大分子水解,把纖維素、半纖維素、果膠、淀粉、脂類和蛋白質(zhì)等物質(zhì),經(jīng)水解發(fā)酵生成水溶性糖、醇、酸等分子較小的化合物;然后,微生物進(jìn)一步把較小分子的化合物降解形成產(chǎn)甲烷菌可直接利用的底物,主要是乙酸鹽、H2和CO2等;最后,產(chǎn)甲烷菌才能利用極簡單的小分子化合物代謝產(chǎn)生CH4和CO2。有機(jī)物質(zhì)在經(jīng)過厭氧生物轉(zhuǎn)化為甲烷的過程中,涉及一系列微生物菌群代謝,它們的底物和特性完全不同。整個(gè)轉(zhuǎn)化過程可以描述為許多種不同微生物類群的直接和間接的共生聯(lián)合關(guān)系。確定的9個(gè)步驟分別由專門的菌群調(diào)節(jié)代謝,這9個(gè)步驟分別為:a.有機(jī)聚合物水解成有機(jī)單體,如糖、有機(jī)酸和氨基酸等;b.有機(jī)單體轉(zhuǎn)化成氫、重碳酸鹽、乙酸、丙酸和丁酸以及其他有機(jī)產(chǎn)物如乙醇、乳酸等;c.產(chǎn)氫產(chǎn)乙酸菌對(duì)還原性有機(jī)物具有氧化作用,能生成氫氣、重碳酸鹽和乙酸;d.同型乙酸菌對(duì)重碳酸鹽的產(chǎn)乙酸呼吸作用;e.硝酸鹽還原菌(NRB)和硫酸鹽還原菌(SRB)對(duì)還原性有機(jī)物的氧化作用生成重碳酸鹽硼乙酸;f.NRB和SRB對(duì)乙酸的氧化作用,生成重碳酸鹽;g.NRB和SRB對(duì)氫氣的氧化作用;h.裂解乙酸的甲烷發(fā)酵;i.重碳酸鹽的產(chǎn)甲烷呼吸作用。 在厭氧消化過程中,微生物的相互作用實(shí)質(zhì)上是各類微生物在代謝上的相互影響。每一種微生物都有其自身的代謝途徑,它們可以在特定的環(huán)境中單獨(dú)行使其代謝功能。因此,研究它們的代謝條件和代謝途徑是非常有用的。但在厭氧消化反應(yīng)器這樣一個(gè)雜居的環(huán)境中,各類型的微生物生活在一起,各自進(jìn)行自身特有的代謝,并形成各自的代謝產(chǎn)物,彼此之間的相互作用,使它們能正常進(jìn)行生命活動(dòng),然而它們所形成的代謝產(chǎn)物可能引起如下后果:a.產(chǎn)物累積引起的反饋抑制效應(yīng);b.代謝產(chǎn)物被其他菌群利用,促進(jìn)其生長;c.抑制其他菌群;d.造成有利或不利于其他菌群的生態(tài)環(huán)境。 這些問題在厭氧消化反應(yīng)器中都有可能出現(xiàn),而在厭氧消化正常運(yùn)行或運(yùn)行失敗時(shí),代謝產(chǎn)物的種類及其濃度都有很大的差別,代謝產(chǎn)物引發(fā)的問題和后果也完全不同。在正常的厭氧消化過程中,各種微生物代謝過程發(fā)生相互偶聯(lián),產(chǎn)物得到協(xié)調(diào),整個(gè)消化過程達(dá)到平衡。相反,任何一個(gè)代謝過程的失調(diào)都有可能引發(fā)整個(gè)過程的破壞,使厭氧消化過程失衡。因此,研究厭氧消化微生物的相互關(guān)聯(lián)性是一個(gè)重要且復(fù)雜的內(nèi)容。 (3)甲烷發(fā)酵過程微生物的代謝關(guān)聯(lián)性 對(duì)厭氧消化過程中發(fā)酵液和氣體成分進(jìn)行分析,特別是將一些功能菌的代謝產(chǎn)物進(jìn)行分析,就不難發(fā)現(xiàn)微生物之間的代謝關(guān)系。 1)水解酸化菌與產(chǎn)甲烷菌之間的相互關(guān)系 下圖所示為某反應(yīng)器內(nèi)污泥厭氧消化過程中揮發(fā)酸和日沼氣產(chǎn)量的關(guān)系。在厭氧消化開始后的初期階段,反應(yīng)器內(nèi)大量產(chǎn)酸,揮發(fā)酸的濃度最高可達(dá)4000mg/L,此時(shí)反應(yīng)器內(nèi)以水解酸化過程為主,此后,反應(yīng)過程逐漸過渡到產(chǎn)甲烷階段。反應(yīng)器內(nèi)水解酸化階段和產(chǎn)甲烷階段是兩個(gè)無法在絕對(duì)意義上分開的過程,這主要是由產(chǎn)酸過程和產(chǎn)甲烷過程沒有達(dá)到代謝平衡所致。由于產(chǎn)酸菌的繁殖速率較高,在厭氧消化開始初期就大量繁殖,產(chǎn)生較大量的揮發(fā)酸,而產(chǎn)甲烷菌的繁殖速率較慢,因此,水解酸化細(xì)菌代謝產(chǎn)生的揮發(fā)酸不能及時(shí)地被產(chǎn)甲烷菌利用,故此階段主要表現(xiàn)為揮發(fā)酸的積累。當(dāng)產(chǎn)甲烷菌數(shù)量增加、微生物區(qū)系相對(duì)完善后,揮發(fā)酸很快被消耗,產(chǎn)甲烷則成為主要特征。由于在揮發(fā)酸增強(qiáng)的同時(shí),產(chǎn)甲烷菌的數(shù)量不斷增加,沼氣產(chǎn)量也隨之增加,此時(shí)水解酸化階段和產(chǎn)甲烷階段構(gòu)成一個(gè)連續(xù)的過程。因此可以采用增加接種物的辦法縮短這兩個(gè)階段之間的時(shí)間差,使產(chǎn)甲烷階段提前。研究人員曾在污泥厭氧消化試驗(yàn)中,在反應(yīng)器內(nèi)接種容積量35%的厭氧消化液作為啟動(dòng)微生物,此時(shí)反應(yīng)器在第2日即可正常產(chǎn)氣,第4日即可達(dá)到產(chǎn)氣高峰,而接種反應(yīng)器容積量15%的厭氧消化液時(shí),產(chǎn)氣高峰出現(xiàn)在第9日,這說明增加接種物的用量可以較好地保證反應(yīng)器的正常啟動(dòng),也說明水解酸化階段和產(chǎn)甲烷階段是可以人為調(diào)控的。
2)水解酸化菌之間相互關(guān)系 在污泥厭氧消化過程中,運(yùn)行正常的消化罐內(nèi)揮發(fā)酸的乙酸濃度最高,為200~1500mg/L;丙酸濃度較低,為300~1000mg/L;丁酸濃度最低,在100mg/L以下,有時(shí)其濃度低于檢測線。這幾種揮發(fā)酸中乙酸的濃度波動(dòng)是最明顯的。當(dāng)乙酸濃度降到比丙酸低的時(shí)候,特別是乙酸濃度低于200mg/L時(shí),丙酸、丁酸的濃度均有明顯的下降。在中溫發(fā)酵過程中,揮發(fā)酸的變化趨勢(shì)也較為近似。上述這些結(jié)果表明,在厭氧消化過程中,產(chǎn)酸菌主要以產(chǎn)乙酸菌為主,產(chǎn)丙酸菌次之,產(chǎn)丁酸菌最少。丙酸、丁酸不能被產(chǎn)甲烷菌直接利用,它們?cè)诒贿M(jìn)一步降解為乙酸后才能轉(zhuǎn)化為甲烷。因此,丙酸和丁酸濃度的下降顯然說明了與產(chǎn)乙酸菌的密切關(guān)系,這充分說明它們之間有相互依賴的關(guān)系和種群消長關(guān)系,這種關(guān)系無論在常溫還是中溫消化過程中都比較穩(wěn)定。 3)產(chǎn)氫和產(chǎn)甲烷菌的相互關(guān)系 自20世紀(jì)80年代奧氏甲烷桿菌被分離后,產(chǎn)氫菌和產(chǎn)甲烷菌之間的共生關(guān)系才為人所知。此后,這一關(guān)系又為其他許多學(xué)者所證實(shí)。在餐廚垃圾與污泥的混合消化過程中也發(fā)現(xiàn)這一現(xiàn)象[2]。在污泥的厭氧消化過程中,當(dāng)產(chǎn)甲烷過程進(jìn)行正常(甲烷濃度60%~70%)時(shí),向反應(yīng)器中加入反應(yīng)器容積20%的餐廚垃圾,當(dāng)pH值降低至6.0時(shí),氫氣濃度出現(xiàn)明顯上升,當(dāng)pH值降低至5.0時(shí),氫氣濃度達(dá)到最大,濃度峰值為50%~55%。氫氣濃度在達(dá)到峰值后出現(xiàn)下降的現(xiàn)象,此時(shí)pH值也隨之上升,甲烷濃度也逐步上升。由此可看出,產(chǎn)氫微生物和產(chǎn)甲烷菌兩者間有明顯的共生關(guān)系,沼氣中氫氣濃度的上升和下降反映了厭氧消化過程中產(chǎn)氫菌和產(chǎn)甲烷菌在代謝上的相互偶聯(lián)。當(dāng)反應(yīng)器的環(huán)境更有利于產(chǎn)氫菌繁殖時(shí),產(chǎn)甲烷菌的活性則不斷下降,此時(shí)氫氣濃度最高時(shí)可達(dá)60%。 |
歡迎分享轉(zhuǎn)載→ http://m.avcorse.com/read-750655.html
Copyright ? 2024 有趣生活 All Rights Reserve吉ICP備19000289號(hào)-5 TXT地圖HTML地圖XML地圖